生物矿化在生物界广泛存在,生物矿物的形成过程及其分子机制一直是地球科学与生命科学的交叉前沿,但也是研究的难点。在最近发表于《国家科学评论》的文章中,中国科学院地质与地球物理研究所李金华研究员和潘永信院士带领的国际研究团队,通过比较基因组学和比较表型组学的关联研究,揭示趋磁细菌系统中调控磁小体生物矿化和链组装的关键基因网络,并提出磁小体生物矿化的通用模式图,为未来更深入地基因功能鉴定和生物仿生矿化提供指导框架,也为地质记录中磁小体化石的识别提供理论依据。
趋磁细菌是一类独特的矿化微生物,能在细胞内合成由生物膜包被的、纳米尺寸磁铁矿(Fe3O4)或胶黄铁矿(Fe3S4)晶体颗粒,也叫磁小体。磁小体常常首尾相连,被组装成单链或者多链结构,作为细胞的生物小磁针,使趋磁细菌能在水生环境的有氧-缺氧过渡带中,沿地磁场方向上下穿梭。地质记录中的磁小体化石因为具有单磁畴尺寸而磁性强、晶型和结构独特而易识别,被认为是最理想的剩磁载体和生物标志矿物,有望成为同时开展古地磁、古环境和古微生物综合研究的新材料。
自首次发现六十多年来,磁小体的合成和链组装过程及其分子机制一直是趋磁细菌领域的研究热点。然而,趋磁细菌在实验室环境下极难生长,依靠纯培养的可控遗传操作系统极难建立。目前,系统的磁小体合成过程及其分子机制仅仅在两株模式菌株AMB-1和MSR-1中展开。这两株趋磁细菌都隶属于假单胞菌门α-变形菌纲的趋磁螺菌属,均只能在细胞内合成单条链的立方八面体形磁铁矿磁小体。大量研究表明,环境中还存在大量其它门类的趋磁细菌,它们的磁小体形貌和链组装模式也多种多样。如何建立不依赖纯培养的研究体系,开展“趋磁细菌类群-磁小体生物矿化”关联研究,是全面认识磁小体合成和链组装机制的关键所在。
为此,研究者首先建立未培养趋磁细菌的比较基因学和比较表型组学研究工作流程。在课题组前期建立的“荧光-电子显微联用”技术的基础上,本次研究引入了宏基因组学和生物信息技术,建立了一套能够在单细胞水平上鉴定趋磁细菌类群和研究磁小体形貌结构特征、获取未培养趋磁细菌的基因组并识别其磁小体合成相关基因(简称磁小体基因)的工作流程图(图1)。通过该工作流程图,本次研究从自然环境中获得了15个未培养趋磁细菌的“细菌类群-磁小体形貌-磁小体岛基因”相关联的数据集,与所有前人报道的32个含有相对完整信息的趋磁细菌数据集进行整合,通过系统地比较基因组学和比较表型组学研究,获得控制磁小体合成和链组装的关键基因,并结合前人在可培养趋磁细菌中的研究成果,建立磁小体生物矿化通用模型。

图1 未培养趋磁细菌基因组和表型组研究的工作流程
通过对47个趋磁细菌数据集的比对分析,建立“趋磁细菌类群-磁小体基因”的相关性(图2)。结果发现:(1)8个mam基因(即mamABEIKMPQ)在所有调研的趋磁细菌基因组中都严格保守,它们都是控制趋磁细菌中磁小体生物合成的普遍过程(磁小体膜形成,铁吸收,磁铁矿成核和磁小体链组装)的核心基因,命名为磁小体核心基因(MACGs);(2)4个mam基因(即mamHFST)只存在于假单胞菌门趋磁细菌中(简称MACGPs);(3)部分mad基因和man基因分别只在脱硫菌门和硝化螺菌门中保守。这表明磁小体基因的含量和排列在不同门类中差异很大,而在同一门类中相对保守。意味着磁小体基因簇的多样性可能是导致不同趋磁细菌类群中磁小体形貌和链结构差异的原因。

图2 47个趋磁细菌数据集的系统发育(a)、基因组完成度(b)、磁小体基因(c)和代表性细胞和磁小体透射电镜图片(d)
通过磁小体“形貌-基因”的关联分析和蛋白质功能的生物信息学预测,识别调控磁小体形貌的关键基因(图3)。结果发现:(1)mms6基因在合成立方八面体的趋磁细菌中高度保守,而在脱硫菌门和硝化螺菌门中缺失,这进一步支持了前人mms6基因控制立方八面体形貌的观点;(2)在假单胞菌门趋磁细菌磁小体基因簇中发现了一个与Mms6蛋白三维结构高度相似的蛋白,命名为Mms6-like,其可能与Mms6具有相似的功能;(3)蛋白保守结构域预测表明,除了Mad10、Mad11蛋白之外,Mad3-5、Mad8以及Mad19也具有金属离子亲和区,可能与直子弹头形磁小体的形貌控制有关;(4)Man-1、-3和-4蛋白同样具有金属离子亲和区,而且仅在硝化螺菌门中高度保守,这说明它们可能控制弯曲的子弹头形磁小体的形貌。

图3 磁小体磁铁矿晶体形貌与磁小体基因相关性
通过磁小体“链结构-基因” 的关联分析和蛋白质功能的生物信息学预测,识别调控磁小体链组装的关键基因(图4)。结果表明:(1)前人鉴定的磁小体链骨架基因mamK在所有趋磁细菌中严格保守,这进一步确认了其在磁小体链组装过程中的关键作用;(2)前人发现的与链结构有关的基因,如mamJ、mamY、mad28以及macAB,只在部分趋磁细菌类群中分布,说明它们在不同趋磁细菌系统发育群中导致了磁小体链结构的多样性;(3)mamK基因的多拷贝导致假单胞菌门暂定η-变形菌纲趋磁细菌的磁小体链由一条变成多条,而多拷贝mamK基因之间的相似性决定了其链结构为双链、四链还是不成链;(4)mcaA基因只有位于磁小体基因簇中时才能发挥在成熟磁小体之间加入新的磁小体的功能;(5)γ-变形菌纲趋磁细菌两个相邻的mamK基因可能受一个操纵子调控,合成一条长的磁小体链;(6)mad24和man5基因的编码蛋白都具有一个SMC结构域和两端的金属离子亲和区,这暗示它们可能分别与脱硫菌门和硝化螺菌门中磁小体链束结构有关。
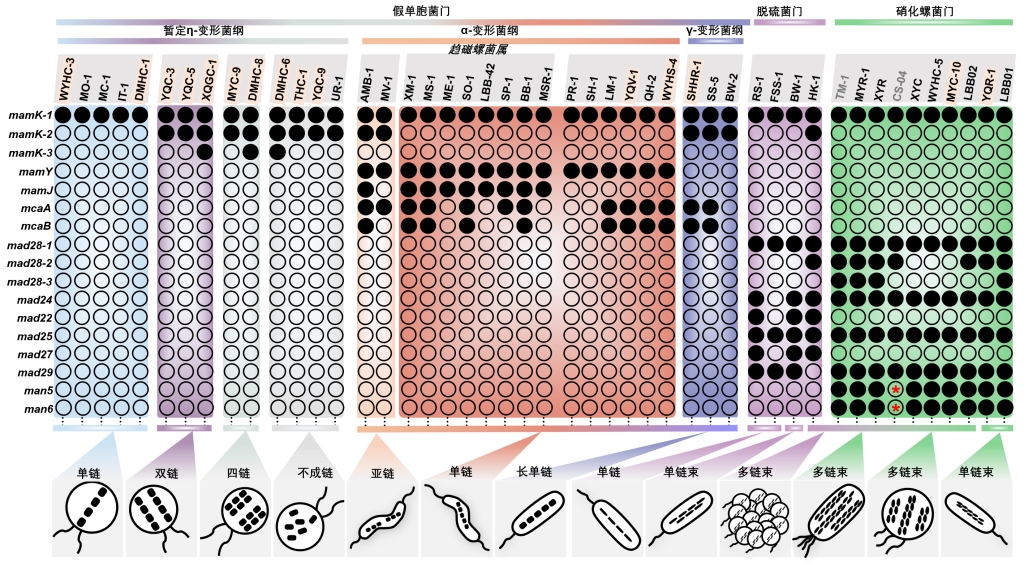
图4 磁小体链结构与磁小体基因相关性
基于上述研究和分析,研究者提出了磁小体合成和链组装的关键基因网络及其通用模式图(图5)。该模型涵盖“假单胞菌门、脱硫菌门和硝化螺菌门”三大门类趋磁细菌,可用于解释“八面体、立方八面体、棱柱形、不弯曲子弹头形和弯曲子弹头形”磁小体合成过程及其分子机制,也可用于解释“单链(束)、多链(束)、非线性链”磁小体组装过程及其分子机制。因此,在未来的研究中,研究者可以借助这个基因网络图,来指导磁小体基因点突变或异源磁小体基因表达实验,进而更好地理解棱柱状和子弹头形磁小体的生物合成和链组装的分子机制。其次,本研究成果为前期课题组建立的“磁小体形貌-趋磁细菌类群”相关性模型,提供了基因证据,这为地质记录中磁小体化石的识别及其古趋磁细菌类群反演、古地磁和古环境信息解译提供理论依据。最后,本研究还揭示,有些蛋白质(如Mad3和Man1)有望通过异源表达,生物仿生矿化合成高度拉长的纳米磁铁矿颗粒,具有比球形或立方八面体磁铁矿颗粒更高的矫顽力,使其更适合于纳米医学和纳米技术的应用。
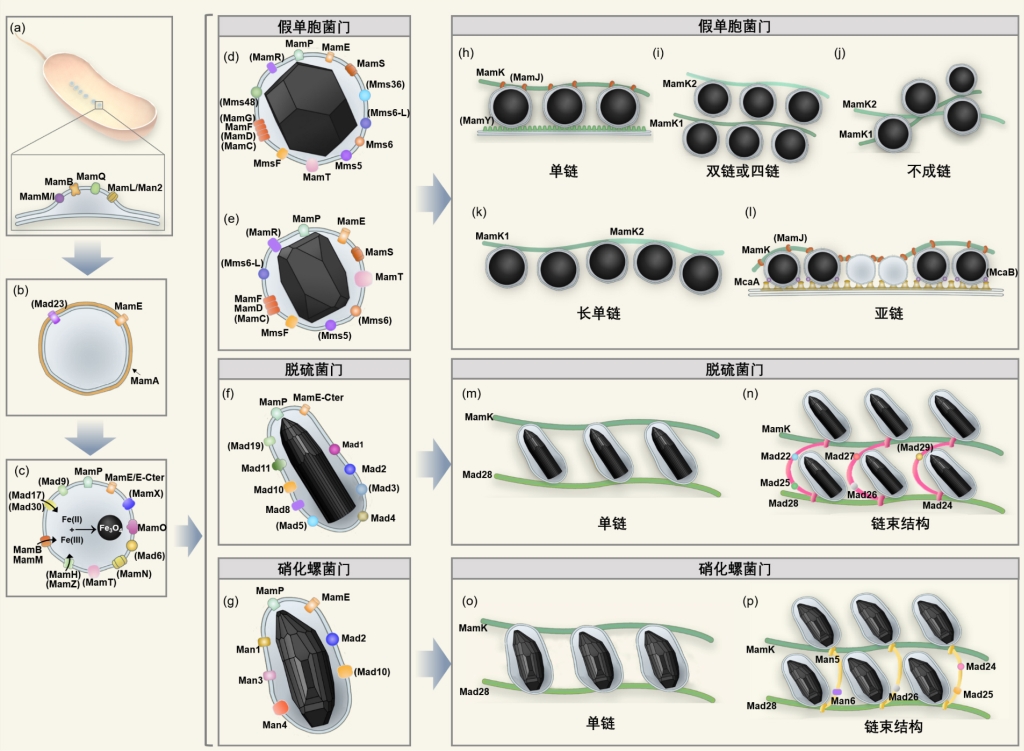
图5 磁小体生物矿化的基因网络及其通用模型
研究成果发表于NSR(刘沛余,郑越,张荣荣,白金伶,朱珂磊,Benzerara Karim,Menguy Nicolas,赵翔,Roberts Andrew,潘永信,李金华*. Key gene networks that control magnetosome biomineralization in magnetotactic bacteria[J]. National Science Review, 2022: nwac238. DOI: 10.1093/nsr/nwac238)。该研究得到了国家自然科学基金(41920104009,41890843和41621004)的资助。